
Page 312 - 第九版生物化学
P. 312
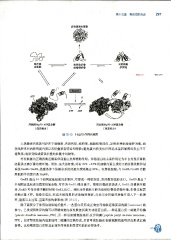
`待折叠 蕃 第十五章蛋白质的合成 297
多肤链 ^空
癹 kky, 2 018
未完全折叠
或错误折叠 @
\/ \ 安 kky,2018
恤广矗,l ` 上p
一
i
气
了
ATP ADP
开放的Hsp70-ATP复合物 闭合的Hsp70-AD吵给物
(低亲和力) (高亲和力)
图 15 -10 Hsp70 作用示意图
人热激蛋白家族可存在于细胞质、内质网腔、线粒体、细胞核等部位,发挥多种细胞保护功能,如
使线粒体和内质网蛋白质以未折叠状态转运和跨膜;避免蛋白质变性后因疏水基团暴露而发生不可
逆聚集;清除变性或错误折叠的肤链中间物等。
有些肤链的正确折叠还需要伴侣蛋白发挥辅助作用 。 伴侣蛋白的主要作用是为非自发性折叠肤
链提供正确折叠的微环境 。 例如,在大肠杆菌,约有 10% -15% 的细胞内蛋白质的正确折叠依赖伴侣
系统 GroEL/GroES, 热激条件下依赖该系统的蛋白质则高达 30% 。 在真核细胞,与 GroEL/ GroES 功能
类似的伴侣蛋白是 Hsp60 。
GroEL 是由 14 个相同亚基组成的多聚体,可形成一桶状空腔,顶部是空腔的出口 。 GroES 是由 7
个相同亚基组成的圆顶状复合物,可作为 GroEL 桶的盖子。需要折叠的肤链进入 GroEL 的桶状空腔
后, GroES 可作为盖子瞬时封闭 GroEL 出口 。 封闭后的桶状空腔为肤链折叠提供微环境,折叠过程需
消耗大量 ATP 。 折叠完成后,形成天然构象的多肤链被释放,尚未完全折叠的肤链可进入下一轮循
环,重复以上过程,直至天然构象形成(图 15-11) 。
除了需要分子伴侣协助肤链折叠外,一些蛋白质形成正确空间构象还需要异构酶 (isomerase) 的
参与。已发现两种异构酶可以帮助细胞内新生肤链折叠为功能蛋白质,一种是蛋白质二硫键异构酶
(protein disulfide isomerase, PDI) , 另一种是肤脯氨酰基顺-反异构酶 (peptide prolyl cis-trans isomerase,
PPI) 。 前者帮助肤链内或肤链间二硫键的正确形成,后者可使肤链在各脯氨酸残基弯折处形成正确
折叠。这些都是蛋白质形成正确空间构象和发挥功能的必要条件。