
Page 309 - 第九版生物化学
P. 309
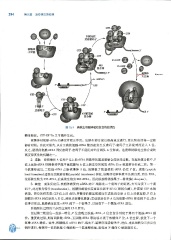
294 第三篇 遗传信息的传递
核糖体
小亚基
I1
安 kky,2018
s ' h~
43S前起始 5' 卢 3'
复合物
图 15-7 真核生物翻译起始复合物的装配
糖体释放 。 GTP-EF-Tu 又可循环生成 。
核糖体对氨酰 -tRNA 的进位有校正作用 。 肤链生物合成以很高速度进行,延长阶段的每一过程
都有时限。在此时限内,只有正确的氨酰 -tRNA 能迅速发生反密码子-密码子互补配对而进入 A 位 。
反之,错误的氨酰-tRNA 因反密码子-密码子不能配对结合而从 A 位解离 。 这是维持肤链生物合成的
高度保真性的机制之一 。
2. 成肤 指核糖体 A 位和 P 位上的 tRNA 所携带的氨基酸缩合成肤的过程 。 在起始复合物中, P
位上起始 tRNA 所携带的甲酰甲硫氨酸与 A 位上新进位的氨酰 tRNA 的 ex- 氨基缩合形成二肤 。 第 一
个肤键形成后,二肤酰 -tRNA 占据核糖体 A 位,而卸载了氨基酸的 tRNA 仍在 P 位 。 成肤 (peptide
bond formation) 过程由肤酰转移酶 (peptidyl transferase) 催化,该酶的化学本质不是蛋白质,而是 RNA,
在原核生物为 23S rRNA, 在真核生物为 28S rRNA 。 因此肤酰转移酶属于一种核酶 (ribozyme) 。
3. 转位 成肤反应后,核糖体需要向 mRNA 的 3 I -端移动一个密码子的距离,方可阅读下一个密
码子,此过程为转位 (translocation) 。核糖体的转位需要延长因子 EF-G( 即转位酶),并需要 GTP 水解
供能 。 转位的结果是:心P 位上的 tRNA 所携带的氨基酸或肤在成肤后交给 A 位上的氨基酸, P 位上
卸载的 tRNA 转位后进入 E 位,然后从核糖体脱落;@成肤后位千 A 位的肤酰 -tRNA 移动到 P 位; @A
位得以空出,且准确定位在 mRNA 的下一个密码子,以接受下一个氨酰 -tRNA 进位 。
肤链延长过程的三步反应如图 15-8 所示 。
经过第二轮进位一成肤—转位, P 位出现 三肤酰 -tRNA ,A 位空留并对应于第四个氨酰-tRNA 进
位 。 重复此过程,则有四肤酰 -tRNA 、五肤酰 -tRNA 等陆续出现千核糖体 P 位, A 位空留,接受下一个
氨酰 -tRNA 进位 。 这样,核糖体从 mRNA 的 5'-端向 3'-端顺序阅读密码子,进位、成肤和转位三步反应
也 循环进行,每循环一次向肤链 C-端添加一个氨基酸残基,肤链由 N-端向 C-端逐渐延长 。