
Page 283 - 第九版生物化学
P. 283
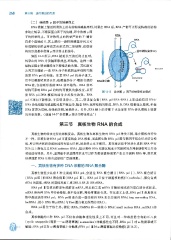
268 第 三 篇遗传信息的传递
( 二 )非依赖 p 因子的转录终止
DNA 模板上靠近转录终止处有些特殊碱基序列,转录出 RNA 后, RNA 产物可以形成特殊的结构
来终止转录,不需要蛋白因子的协助,即非依赖 p 因 ,-?DNA
子的转录终止 。 可导致终止的转录产物的 3'-端常
有多个连续的 u, 其上游的 一 段特殊碱基序列又可 5 ' 3一'
形成鼓糙状的茎环或发夹形式的二级结构,这些结
构的形成是非依赖 p 因子终止的信号 。 茎环结构(富含GC)
如图 14-8 所示, RNA 链延长至接近终止区时,
转录出的 RNA 片段随即形成茎-环结构 。 这种二级
结构是阻止转录继续向下游推进的关键。其机制可 癹 kk y, 201 8
从两方面理解:一是 RNA 分子形成的茎环结构可能
改变 RNA pol 的构象 。 注意 RNA pol 的分子量大,
它不但覆盖转录延长区,也覆盖部分 3' -端新合成的 RNA 聚合酶 3'
RNA 链,包括刚形成的 RNA 茎环结构 。 RNA 茎环
结构可影响 RNA pol 的结构而使其构象改变,从而 3'5'
使 RNA pol-DNA 模板的结合方式发生改变 。 RNA 图 14-8 非依赖 p 因子的转录终止模式
pol 不再向下游移动,于是转录停止 。 其二,转录复合物 (RNA pol-DNA-RNA) 上形成的局部 RNA/
DNA 杂化短链的碱基配对是不稳定的,随着 RNA 茎环结构的形成, RNA 从 DNA 模板链上脱离,单链
DNA 复原为双链,转录泡关闭,转录终止 。 另外, RNA 链上的多聚 U 也是促使 RNA 链从模板上脱落
的重要因素 。 (动画 14 -3" 非依赖 p 因子的转录终止")
第三节 真 核生物 RNA 的合成
真核生物的转录过程比原核复杂 。 真核生物和原核生物的 RNA pol 种类不同,结合模板的特性
不一样 。 原核生物 RNA pol 可直接结合 DNA 模板,而真核生物 RNA pol 需与辅因子结合后才结合模
板,所以两者的转录起始过程有较大区别,转录终止也不相同 。 真核基因组中转录生成的 RNA 中有
20% 以上存在反义 RNA(antisense RNA), 提示某些 DNA 双链区域在不同的时间点两条链都可以作为
模板进行转录 。 另外,基因组中的基因间区也可以作为模板被转录而产生长非编码 RNA 等 , 提示真
核基因组 RNA 生物合成是很广泛的现象 。
—、真核生物有多种 DNA 依赖的 RNA 聚合酶
真核生物至少具有 3 种主要的 RNA pol, 分别是 RNA 聚合酶 I (RNA pol I) 、 RNA 聚合酶 II
(RNA pol II) 和 RNA 聚合酶皿 (RNA pol ill) 。 RNA pol I 位千细胞核的核仁 (nucleolus) , 催化合成
rRNA 的前体, rRNA 的前体再加工成 28S 、5 . 8S 及 18S rRNA 。
RNA pol II 在核内转录生成前体 mRNA, 然后加工成 mRNA 并输送给胞质的蛋白质合成体系 。
mRNA 是各种 RNA 中寿命最短、最不稳定的,需经常重新合成 。 在此意义上说, RNA pol II 是真核生
物中最活跃的 RNA pol 。 RNA pol II 也合成一些非编码 RNA 如长非编码 RNA (long non-coding RNA,
lncRNA) 、微小 RNA 和 piRNA( 与 Piwi 蛋白相互作用的 RNA) 。
RNA pol 皿位于核仁外,催化 tRNA 、 5SrRNA 和一些核小 RN A (small nuclear RN A , snRNA) 的
合成 。
真核细胞的三种 RNA pol 不仅在功能和理化性质上不同,而且对一种毒蘑菇含有的环八肤
(cyclic octapeptide) 毒素 -a-鹅膏簟碱 (a -amanitine) 的敏感性也不同, RNA pol I 对 a-鹅 膏 簟碱不
炉, 敏感; RNA pol II 对 a-鹅膏覃碱十分敏感; RNA pol 皿对 a -鹅膏覃碱比较敏感(表 14-2) 。